
„Prochem jesteś i w proch się obrócisz”.
Ten werset z Księgi Rodzaju 3 bardzo dobrze oddaje los pięknie zaprojektowanych organizmów po wejściu grzechu na świat.
Ale ile czasu zajmuje to, aby cząsteczki organiczne, z których jesteśmy zbudowani, rozpadły się po śmierci?
Ogólnie rzecz biorąc, im więcej czasu upłynęło od śmierci, tym większy stopień rozkładu powinno się obserwować. Jest to prawdą szczególnie w przypadku tkanek miękkich – części organizmu, które nie są zmineralizowane (takich jak skóra, mięśnie lub naczynia krwionośne).
Mary Schweitzer i jej badania
W 1993 r. Mary Schweitzer związana wówczas z Muzeum Gór Skalistych, udostępniła dane sugerujące możliwość zachowania tkanek miękkich i biocząsteczek w kości Tyrannosaurus rex żyjącego hipotetycznie 68 milionów lat temu1.
Jej odkrycia spotkały się z dużym sprzeciwem i sceptycyzmem. Podobne obserwacje naczyń krwionośnych, kolagenu i osteocytów z kości dinozaura były publikowane przez Romana Pawlickiego i jego współpracowników od 1966 r.2, ale nie wywołały zbyt wielu dyskusji, prawdopodobnie dlatego, że „Park Jurajski”, który spopularyzował ten temat, nie został jeszcze napisany i nakręcony.
W ciągu ostatnich dwóch dziesięcioleci Mary Schweitzer i jej grupa znaleźli dodatkowe przykłady i wykorzystywali coraz szerszy wachlarz technik analitycznych do udokumentowania swoich odkryć3-9. W związku z tym możliwość zachowania oryginalnej tkanki miękkiej dinozaura i biocząsteczek staje się coraz bardziej akceptowana, a ten post na blogu analizuje pewne kwestie spośród tych, które zostały opublikowane na ten temat w ciągu ostatniego półtora roku.
Okazy dinozaurów z zachowanymi tkankami miękkimi zgłoszone w latach 2016–2017
Dinozaury i ich trzy różne szkielety – w literaturze opisano – z kośćmi oddzielonymi od siebie lub połączonymi, ze szczególnym uwzględnieniem lub podkreśleniem obecności tkanek miękkich.
Poznajmy dinozaury
Ceratops ptasiomiedniczy
Pierwszy, ceratops ptasiomiedniczy (Psittacosaurus sp.) z dolnej kredy w Chinach (ryc. 1), ma skórę zachowaną w postaci ściśniętej cienkiej powłoki z charakterystycznymi wzorami pigmentacyjnymi10. Uważa się, że pigmenty reprezentują oryginalną materię organiczną, a dokładniej pozostałości melaniny. Ta interpretacja została oparta na mikroskopii SEM, ukazującej jajowate wzory podobne do melanosomów (organelli zawierających melaninę).

Ankylozaur
Drugi, ankylozaur (Zuul crurivastator) z górnej kredy w Montanie, zachowuje struktury powłokowe, takie jak osteodermy, z ciemnymi otoczkami prawdopodobnie składającymi się z oryginalnej keratyny11. Artykuł opisujący skamieliny nie przedstawia chemicznej ani mikroskopowej analizy tkanki miękkiej, ale wymienia ją jako przedmiot dalszych badań.
Ankylozaur (Borealopelta markmitchelli)
Trzeci szkielet dinozaurów to także ankylozaur (Borealopelta markmitchelli) z dolnej kredy w Albercie12. Ten przegubowy szkielet znaleziono w osadach morskich, w formacji, w której znaleziono ichtiozaury i plezjozaury, ale nigdy dinozaury.
Osady otaczające wykazują oznaki szybkiego pochówku ciała zwierzęcia, przy braku śladów grzebania, pomimo kilku nor w okolicznych złożach. Wyjątkowy poziom zachowania skamielin obejmuje poziom molekularny z resztkami materii organicznej w łuskach i osłonach rogów pancerza.
Dinozaury – analiza chemiczna
Analiza chemiczna za pomocą spektroskopii masowej wykazała obecność melaniny w pozostałości organicznej, zwłaszcza feomelaniny (czerwonawo-brązowy pigment). Wydaje się, że melanosomy nie zostały zachowane w tym osobniku.
Być może mniej imponujące niż zachowanie tkanek miękkich, ale równie interesujące, są dowody zachowania w tkance kostnej dinozaura oryginalnych składników molekularnych o wyraźnym składzie chemicznym. Ta sygnatura chemiczna została wykorzystana do udowodnienia obecności kości rdzeniastej (rodzaj kości produkowanej przez dojrzałe samice ptaków podczas owulacji) w szczątkach T. rex z górnej kredy w Montanie13.
Mark Armitage dostarczył nową dokumentację14 na temat znakomicie zachowanych naczyń krwionośnych, osteocytów i kolagenu na poziomie submikronowym w rogu i kościach żeber Triceratops horridus, które odkrył w złożach górnej kredy w Montanie15. Obserwacje oparte są na mikroskopii i głównie z rozpuszczonego materiału kostnego. Kolejnym krokiem w tym projekcie powinna być analiza chemiczna materiału. Ten przykład zachowania tkanek miękkich jest szczególnie oszałamiający, biorąc pod uwagę stosunkowo silne zwietrzenie rogu (znaleziono korzenie, strzępki grzybów i szczątki owadów przemieszczające się przez róg).
Mary Schweitzer i współpracownicy opublikowali nowe badanie16 dotyczące szczątków hadrozaura Brachylophosaurus canadensis (górna kreda w Montanie), które wcześniej dostarczyły dowodów na zachowanie endogennych biocząsteczek6. Używając bardziej rygorystycznego protokołu przygotowania próbek i technik spektrometrii masowej o wyższej rozdzielczości, odzyskali osiem sekwencji peptydowych kolagenu z kości hadrozaura. Dwie spośród zidentyfikowanych sekwencji były repliką niektórych znalezionych w poprzednim badaniu, podczas gdy pozostałe sześć było nowe.
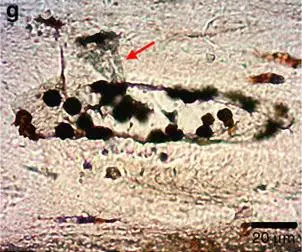
Po ważnym badaniu zaprezentowano wyniki uzyskane z dinozaura zauropodomorficznego (Lufengosaurus) z dolnej jury w Chinach17. Płaskie, przezroczyste fragmenty tkanki miękkiej usytuowane wzdłuż i wewnątrz kanałów naczyniowych w kości żebrowej (ryc. 2) analizowano spektroskopią w podczerwieni stosowaną bezpośrednio in situ, a nie na przetworzonych próbkach kości. Ta nieniszcząca technika zapobiega możliwości zanieczyszczenia próbki podczas rozpuszczania. Obserwowane widmo absorpcyjne było charakterystyczne i typowe dla kolagenu. Ponadto w kanałach naczyniowych (ryc. 2) i lukach pozostawionych przez osteocyty znaleziono cząsteczki hematytu (tlenku żelaza). Cząsteczki interpretowano jako tworzące się z jonów żelaza przyłączonych do komórek krwi i białek wiążących żelazo. Według autorów badanie to dostarczyło „niezaprzeczalnych, wyraźnych dowodów na to, że pozostałości kolagenu i białka zostały zachowane wewnątrz centralnych kanałów naczyniowych osteonów tego wczesnego dinozaura”.
Dinozaury w świetle najnowszych odkryć naukowych
Wreszcie, intrygujący abstrakt został zaprezentowany na spotkaniu Canadian Society of Vertebrate Paleontology w 2017 r.18 Skamieliny z formacji Dinosaur Park Formation z górnej kredy w Albercie wykazały nieoczekiwanie wysoki stopień zachowania tkanek miękkich. Zbiór próbek kości z 25 osobników (w tym 16 z dinozaurów), reprezentujących różny stopień rozczłonkowania i zachowanych zarówno w piaskowcu jak i w mule, rozpuszczono i przebadano pod kątem zachowania tkanek miękkich. Spośród 22 próbek, które się pomyślnie rozpuściły, 20 (w tym próbki z dinozaurów) dało wynik pozytywny jeśli chodzi o tkanki miękkie. Wygląda na to, że zachowanie tkanek miękkich w Dinosaur Park Formation może być bardziej powszechne niż oczekiwano, niezależnie od rodzaju osadu lub stopnia rozczłonkowania osobnika.
Opracowania omawiające proces konserwacji
Zrozumienie drogi, w której cząsteczki organiczne mogą być zachowane przez dziesiątki do setek milionów lat, jest znaczącym wyzwaniem dla tych, którzy wyznają chronologię „głębokiego czasu”. Na przykład uważa się, że białka ulegają znacznej degradacji w krótszych ramach czasowych wynoszących kilkadziesiąt tysięcy lat19. Dlatego kilka badań próbuje odkryć potencjalne mechanizmy, które mogły być przyczyną wyjątkowego zachowania tkanek miękkich w szczątkach dinozaurów.
Niektórzy sugerują, że być może naczynia krwionośne i struktury podobne do osteocytów w kościach dinozaurów nie reprezentują oryginalnego materiału organicznego, ale są naśladownictwem stworzonym przez bakteryjne biofilmy kolonizujące wgłębienia kości20.
Jednak Schweitzer i inni21 przedstawili dane z rzeczywistych eksperymentów z biofilmami bakteryjnymi, aby odrzucić tę hipotezę jako nieodpowiednią. Co ciekawe, podczas przygotowywania próbek kości do swoich eksperymentów zaobserwowali, że usuwanie substancji organicznych z kości nie jest łatwe, nawet podczas surowej obróbki, w tym powtarzanych cyklów ekstremalnego ciepła, wybielacza i obróbki enzymatycznej. Sugerują, że zamknięte w zbitej kości korowej niestałe substancje organiczne mogą przetrwać dłużej.
W swoim artykule na temat zachowanego kolagenu z kości Lufengosaurus Lee i inni odkryli, że kolagen zachował się tylko w kanałach naczyniowych, a nie w macierzy kostnej17. Biorąc pod uwagę, że wnętrze kanałów naczyniowych często zawierało cząstki hematytu, autorzy zasugerowali, że kolagen został zachowany, ponieważ pozostał uwięziony pomiędzy konkrecjami hematytów wewnątrz naczyń a otaczającymi je węglanowymi minerałami apatytowymi w macierzy kostnej.
Czy realistyczne jest sądzić, że te oryginalne tkanki naprawdę zostały zachowane przez dziesiątki milionów lat? Czy są one raczej dowodem na znacznie krótszy czas, jaki upłynął od śmierci skamieniałego organizmu, rzędu tysięcy lat?
Wreszcie, niektórzy wciąż kwestionują wiarygodność wyników opublikowanych przez Mary Schweitzer i jej grupę. Na przykład Buckley i inni22 wykazali, że do wszystkich opublikowanych przypuszczalnych sekwencji peptydów z dinozaurów T. rex i B. canadensis pasują sekwencje kolagenu z kości strusia. Sugeruje się, że nie można wykluczyć zanieczyszczenia krzyżowego próbek dinozaurów materiałem strusia w laboratorium.
Dinozaury – rozważania końcowe
Dyskusja wokół zachowania tkanek miękkich dinozaurów jest fascynującym przykładem zmiany paradygmatu w nauce. Mimo że wciąż napotykają na pewien opór, dowody na obecność endogennego materiału biomolekularnego w skamielinach doprowadziły do rozprzestrzeniania się nowych obserwacji i otwartości na poszukiwanie danych, które wcześniej były pomijane tylko dlatego, że uważano je za coś poza możliwościami.
Oczywiste jest, że obszar ten ma ogromny potencjał wzrostu, w tym lepszą systematyzację rodzaju cząsteczek, które występują bardziej powszechnie w zapisie kopalnym, oraz możliwych różnych poziomów degradacji i rozkładu obserwowanych w tych biocząsteczkach na różnych poziomach stratygrafii.
Ten obszar badań jest bardzo istotny dla modelu pochodzenia, ponieważ ma on implikacje dla dyskusji na temat długiej kontra krótkiej chronologii życia na ziemi. Dinozaury awangardą zmian w dziedzinie paleontologii?
Czy realistyczne jest sądzić, że te oryginalne tkanki naprawdę zostały zachowane przez dziesiątki milionów lat?
Czy są one raczej dowodem na znacznie krótszy czas, jaki upłynął od śmierci skamieniałego organizmu, rzędu tysięcy lat? Mam wrażenie, że odpowiedź na te pytania w dużej mierze nie będzie zależeć od samych dowodów. Jeśli chodzi o pochodzenie i nauki historyczne, „złote rozwiązanie” lub niepodważalny dowód modelu są zazwyczaj nieuchwytne. Osoby zaangażowane w długą chronologię prawdopodobnie będą próbowały znormalizować coś, co wcześniej uważano za wyjątkowe, prezentując liczne scenariusze tego, w jaki sposób zachowanie tkanek miękkich w „głębokim czasie” jest możliwe.
Być może pozytywnym rezultatem tych wysiłków będzie lepsze zrozumienie struktury biomolekularnej, termodynamiki, dróg rozpadu i interakcji z otaczającym środowiskiem chemicznym. Jednak ci, którzy wyznają chronologię biblijną, również będą mieli okazję wskazać ewentualne niedociągnięcia postulowanych mechanizmów zachowania tkanek miękkich w „głębokim czasie”.
Co więcej, jeśli zachowanie tkanek miękkich okaże się bardziej powszechne niż wcześniej sądzono, a nie „wyjątkowe”, wówczas taka linia dowodów dobrze pasowałaby również do krótkiej chronologii i modelu potopu w kontekście pochodzenia, niekoniecznie „dowodząc”, ale z pewnością pozostając w zgodzie z biblijnym światopoglądem. Rzeczywiście najbardziej doskonałe i nieskazitelne przykłady zachowania oryginalnych tkanek miękkich pozostaną prawdopodobnie trudną łamigłówką dla tych, którzy przypisują im wiek obejmujący okresy tak ogromne, że aż trudne do wyobrażenia.
Przypisy
1. Schweitzer, M.H., Biomolecule preservation in Tyrannosaurus rex.Journal of Vertebrate Paleontology, 1993. 13(Supplement to n. 3): p. 56A.
2. Pawlicki, R., A. Korbel, and H. Kubiak, Cells, Collagen Fibrils and Vessels in Dinosaur Bone.Nature, 1966. 211(5049): p. 655-657.
3. Schweitzer, M.H., et al., Soft-tissue vessels and cellular preservation in Tyrannosaurus rex. Science, 2005. 307(5717): p. 1952-1955.
4. Asara, J.M., et al., Protein sequences from Mastodon and Tyrannosaurus rex revealed by mass spectrometry. Science, 2007. 316(5822): p. 280-285.
5. Organ, C.L., et al., Molecular phylogenetics of Mastodon and Tyrannosaurus rex. Science, 2008. 320(5875): p. 499.
6. Schweitzer, M.H., et al., Biomolecular characterization and protein sequences of the Campanian Hadrosaur B. canadensis. Science, 2009. 324(5927): p. 626-631.
7. San Antonio, J.D., et al., Dinosaur Peptides Suggest Mechanisms of Protein Survival. PLoS One, 2011. 6(6): p. e20381.
8. Schweitzer, M.H., et al., Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules. Bone, 2013. 52(1): p. 414-423.
9. Schweitzer, M.H., et al., A role for iron and oxygen chemistry in preserving soft tissues, cells and molecules from deep time. Proceedings of the Royal Society B: Biological Sciences, 2014. 281(1775).
10. Vinther, J., et al., 3D Camouflage in an Ornithischian Dinosaur. Current Biology, 2016. 26(18): p. 2456-2462.
11. Arbour, V.M. and D.C. Evans, A new ankylosaurine dinosaur from the Judith River Formation of Montana, USA, based on an exceptional skeleton with soft tissue preservation. Royal Society Open Science, 2017. 4(5): p. 161086.
12. Brown, C.M., et al., An Exceptionally Preserved Three-Dimensional Armored Dinosaur Reveals Insights into Coloration and Cretaceous Predator-Prey Dynamics. Current Biology, 2017. 27(16): p. 2514-2521e3.
13. Schweitzer, M.H., et al., Chemistry supports the identification of gender-specific reproductive tissue in Tyrannosaurus rex. 2016. 6: p. 23099.
14. Armitage, M.H., Preservation of Triceratops horridus tissue cells from the Hell Creek Formation, MT. Microscopy Today, 2016. 24: p. 18-23.
15. Armitage, M.H. and K.L. Anderson, Soft sheets of fibrillar bone from a fossil of the supraorbital horn of the dinosaur Triceratops horridus. Acta Histochemica, 2013. 115(6): p. 603-608.
16. Schroeter, E.R., et al., Expansion for the Brachylophosaurus canadensis Collagen I Sequence and Additional Evidence of the Preservation of Cretaceous Protein. Journal of Proteome Research, 2017. 16(2): p. 920-932.
17. Lee, Y.-C., et al., Evidence of preserved collagen in an Early Jurassic sauropodomorph dinosaur revealed by synchrotron FTIR microspectroscopy. Nature Comminications, 2017. 8: p. 14220.
18. van der Reest, A.J. and P.J. Currie, Preliminary results of an investigation into the preservation of soft tissue structures in bone from the Dinosaur Park Formation. Vertebrate Anatomy Morphology Palaeontology, 2017. 4: p. 49.
19. Wadsworth, C. and M. Buckley, Proteome degradation in fossils: investigating the longevity of protein survival in ancient bone. Rapid Communications in Mass Spectrometry, 2014. 28: p. 605-615.
20. Kaye, T.G., G. Gaugler, and Z. Sawlowicz, Dinosaurian soft tissues interpreted as bacterial biofilms. PLoS One, 2008. 3(7): p. e2808.
21. Schweitzer, M.H., A.E. Moyer, and W. Zheng, Testing the Hypothesis of Biofilm as a Source for Soft Tissue and Cell-Like Structures Preserved in Dinosaur Bone. PLoS One, 2016. 11(2): p. e0150238.
22. Buckley, M., et al., A fossil protein chimera; difficulties in discriminating dinosaur peptide sequences from modern cross-contamination. Proceedings of the Royal Society B: Biological Sciences, 2017. 284(1855).
Dinozaury – sięgnij po więcej wiedzy
Najnowsze odkrycia sugerują, że niektóre dinozaury składały jaja o miękkiej skorupce – tak jak wiele dzisiejszych gadów (np. węże i żółwie). Czytaj więcej na ten temat. Artykuł w języku angielskim.
