
Wstęp
Są tylko trzy znane grupy latających kręgowców. Obecnie istnieją dwie: ptaki i nietoperze. Trzecia grupa jest całkowicie wymarła i znana tylko ze skamielin: pterozaury. Często nazywane potocznie „pterodaktylami”, pterozaury są różnorodną grupą gadów całkowicie obcych naszym współczesnym umysłom, ale pod pewnymi względami są niewiarygodnie znane – przypominają folklorystyczne smoki. Te prehistoryczne smoki występowały w różnych rozmiarach, od niewielkiej 25-centymetrowej rozpiętości skrzydeł w przypadku Nemicolopterus crypticus (choć uważa się, że nie był to osobnik w pełni dorosły (Wang i in., 2008)) po ogromną rozpiętość skrzydeł azdarchów, takich jak Quetzalcoatlus northropi, wynoszącą 10-11 metrów (Witton i in., 2010), czyniąc te azdarchy największymi znanymi latającymi zwierzętami wszechczasów. Wielkość nie była jedynym znaczącym czynnikiem różnorodności wśród pterozaurów. Niektóre miały długie szyje i krótkie ogony, podczas gdy inne miały krótkie szyje i długie ogony. Niektóre miały wyszukane i dziwaczne grzebienie czaszki, niektóre miały bezzębne szczęki, niektóre były filtratorami, a niektóre miały zęby wieloguzkowe. Ogromna różnorodność pterozaurów jest zdumiewająca, a ich skamieliny pochodzą ze wszystkich siedmiu kontynentów (Barrett i in., 2008), a także z Grenlandii (Jenkins i in., 2001) i Nowej Zelandii (Wiffen i Molnar, 1988). Skamieniałości pterozaurów znajdują się w skałach mezozoicznych, od noryku górnego triasu do mastrychtu górnej kredy (Butler i in., 2009; Unwin, 2006; Wellnhofer, 1991). Zakres ten jest prawie identyczny z zasięgiem dinozaurów, jednak pterozaury nie są dinozaurami. Jedną z pośród wielu cech, których pterozaury nie posiadają, jest perforowana panewka, którą mają dinozaury – otwór w biodrze, który jest miejscem połączenia dla kości udowej. Chociaż określamy pterozaury mianem gadów, nie przypominałyby one żadnego z dzisiaj spotykanych gadów. Z pewnością nie ma wokół nas latających gadów (chociaż są szybujące węże i jaszczurki), ale pterozaury posiadały również pewien rodzaj osłonki składającej się z włoskowatych struktur zwanych pycnofibres (Kellner i in., 2009). Z pewnością byłyby cudownymi do oglądania stworzeniami, majestatycznie szybującymi daleko w górze lub łopoczącymi wzdłuż złotych plaż w poszukiwaniu kąsek zmywanych przez rozbijające się fale.
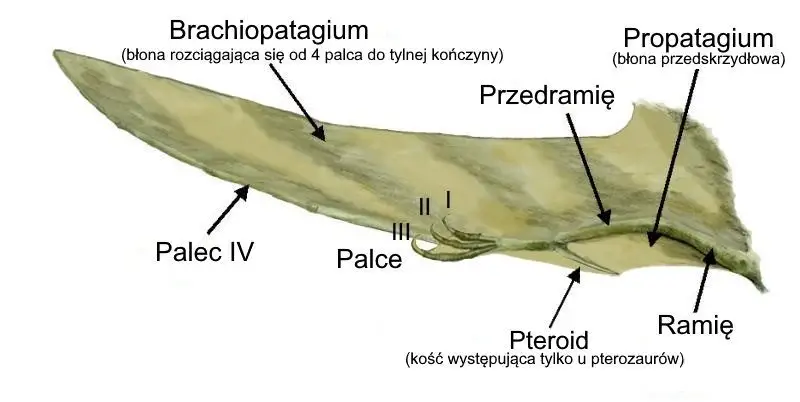
Uznanie wyjątkowości konstrukcji pterozaura jest nieuniknione. W rzeczywistości Kellner (1996) opisał 33 cechy wspólne dla wszystkich pterozaurów, których nie posiadało żadne inne zwierzę. Nawet pobieżne spojrzenie na szkielet pterozaura pozwala rozstrzygnąć, że tak naprawdę nie ma takiego drugiego zwierzęcia. Chociaż istniały trzy grupy kręgowców posiadających zdolność lotu, wszystkie trzy miały radykalnie różne konstrukcje skrzydeł. U pterozaurów skrzydło składa się z błony, która rozciąga się od ciała do kości ramienia (ryc. 1). Czwarty palec u pterozaurów jest w charakterystyczny sposób wydłużony, co pozwala na rozciągnięcie skrzydła znacznie dalej poza zasięg samego ramienia. Szczególną cechą pterozaurów jest również kość o nazwie pteroid, która pozwala błonie zwanej propatagium rozciągać się od nadgarstka z powrotem do ramienia przed główną częścią skrzydła. Na podstawie danych pochodzących ze skamielin wydaje się, że pterozaury miały również odcinek skóry rozciągający się od tylnych nóg do ogona, zwany uropatagium.
Pytanie, które pojawia się w kręgach paleontologicznych po odkryciu takiej wyjątkowej grupy, brzmi: „Skąd pochodzą te stworzenia?” Konwencjonalni paleontolodzy, opierając się wyłącznie na naturalistycznej ewolucji, intensywnie poszukiwali odpowiedzi na to pytanie, przeszukując triasowy zapis skamielin pterozaura. Pomimo ponad 200 lat poszukiwań obraz stał się jeszcze bardziej skomplikowany.
Odkrycie pterozaurów w triasie
Chociaż pierwszą skamielinę pterozaura znaleziono w 1784 r., żadne okazy triasowe nie były znane do 1973 r., kiedy Rocco Zambelli opisał pterozaura Eudimorphodon ranzii (Wellnhofer, 1991) pochodzącego ze skał morskich we Włoszech. Holotyp (okaz typu) E. ranzii (ryc. 2) został dobrze zachowany, ale był również bardzo unikatowy wśród pterozaurów. E. ranzii posiadał zęby wieloguzkowe, w przeciwieństwie do późniejszych pterozaurów, które często posiadały zęby jednoguzkowe. Pamiętaj, że tradycyjnie uważa się, że zęby wieloguzkowe u ssaków wyewoluowały z prostych szpiczastych zębów gadzich. Ciało Eudimorphodon nie było szczególnie niezwykłe w przypadku „rhamphorhynchoidea” („prymitywnych” form o zazwyczaj długich ogonach), chociaż późniejsze odkrycia wykazały, że niektóre triasowe pterozaury nie posiadały usztywniających prętów w ogonach, jak to było w przypadku jurajskich pterozaurów o długich ogonach (Dalla Vecchia, 2002).

Eudimorphodon był z pewnością szczególny, ale nie wyglądał na brakujące ogniwo, na jakie liczyli paleontolodzy. W rzeczywistości wielu badaczy uważa Eudimorphodon za krewnego Campylognathoides, który uważany jest za jednego z bardziej zaawansowanych „rhamphorhynchoidea”. Dalsze odkrycia we Włoszech ujawniły dwa inne pterozaury triasowe: Peteinozaura i Preondaktyla. Peteinozaur uważany jest za przedstawiciela dimorfodontów, podczas gdy Preondaktyl zdaje się przeciwstawiać przypisaniu do określonej grupy. Unwin (2003) uważał Preondaktyla za najbardziej prymitywnego pterozaura, podczas gdy Kellner (2003) uważał anurognaty za najbardziej prymitywną grupę pterozaurów (pomimo faktu, że anurognaty nie pojawiają się aż do środkowej jury). Preondaktyl posiada kilka unikalnych cech, ale ma również wszystkie niezbędne cechy pterozaura.
Na przestrzeni lat odkryte zostały kolejne pterozaury triasowe. We Włoszech znaleziono nowy gatunek Eudimorphodon: E. rosenfeldi, który został później umieszczony w nowym rodzaju (Carniadactylus rosenfeldi) (Dalla Vecchia, 2009a). Inny gatunek Eudimorphodon (E. cromptonellus) został znaleziony w triasowych złożach jeziornych Grenlandii (Jenkins i in., 2001), jednak uważa się, że E. cromptonellus również nie należy do rodzaju Eudimorphodon (Dalla Vecchia, 2009a). Odkryto również Austriadaktyla, czubatą formę z Austrii (Dalla Vecchia i in., 2002), znalezioną później również we Włoszech (Dalla Vecchia, 2009b). Szwajcaria wydała jeszcze dwa triasowe pterozaury: Retikodaktyla i Caviramusa, chociaż Dalla Vecchia (2009a) doszedł do wniosku, że Caviramus to tak naprawdę Retikodaktyl. Istnieje kilka okazów z Alp, które najprawdopodobniej reprezentują nowe gatunki (Dalla Vecchia, 2009a), takie jak BSP 1994 I 51, który został pierwotnie opisany jako okaz E. ranzii (Wellnhofer, 2003). Niedawno odkryta forma z Brazylii, Faxinalipterus, została okrzyknięta najbardziej prymitywnym ze znanych pterozaurów (Bonaparate i in., 2010), ale Dalla Vecchia (2013) doszedł do wniosku, że szczątki najprawdopodobniej nawet nie należą do pterozaura. Uważa się, że niedawno odkryta szczęka z Formacji Chinle Parku Narodowego Skamieniałego Lasu (Petrified National Forest, Arizona, USA) pochodzi od pterozaura (Whatley i in., 2013). W Europie i Ameryce Północnej istnieje kilka odkryć wieloguzkowych zębów przypisywanych pterozaurom z noryku i retyku, ale Dalla Vecchia (2013) zauważył, że niepowiązane z nimi, a pochodzące z tego samego okresu synapsydy – chiniquodonts i dromatheriids – mają bardzo podobne zęby, co poddaje w wątpliwość charakter tych zębów jako należących do pterozaura.
Dyskusja wokół kopalnego zapisu triasowego pterozaura
Z listy tej wynika jedno: na samym początku występowania pterozaurów w zapisie kopalnym istnieje już kilka gatunków, które rozciągają się na znaczną część globu (zwłaszcza gdy uwzględni się obecnie nieopisany okaz z Argentyny). W triasie pojawiają się co najmniej 4 rodziny pterozaurów: Preondactylidae, Campylognathoididae, Dimorphodontidae i Raeticodactylidae (Andres i in., 2014). Zatem najwcześniejsze pojawienie się pterozaurów jest proste. W rzeczywistości różnorodność jest jeszcze bardziej złożona, jako że Dalla Vecchia (2009a) ustalił, że E. cromptonellus oraz okaz BSP 1994 I 51 wydają się znajdować poza Campylognathoididae (chociaż stwierdził, że raeticodactylids zaliczają się do Campylognathoididae). Co ciekawe, jest to podobna sytuacja jak w przypadku innych grup w zapisie kopalnym. Pelykozaury, terapsydy, dinozaury i wiele innych grup pojawiają się w zapisie kopalnym od razu z dużą różnorodnością. Jest to również zgodne ze wzorem obserwowanym w eksplozji kambryjskiej, aczkolwiek na znacznie mniejszą skalę.
Ponieważ pterozaury są tak niewiarygodnie odrębne (tj. są nieciągłe w stosunku do wszystkich innych organizmów), wydaje się mało prawdopodobne, aby wyewoluowały z gadów lądowych
Pomimo dużej różnorodności okazy pterozaurów triasowych są niezwykle rzadkie. Liczba znanych okazów kopalnych określonych pterozaurów triasowych wynosi 32 lub mniej (McLain i in., 2014). Różni się to znacznie od wielu innych zapisów skamielin pterozaurów, gdzie poszczególne gatunki pterozaurów mogą być znane z 30, a nawet z ponad 1000 okazów (Kellner, 2010). Prawie wszystkie znane niekwestionowane okazy pterozaurów pochodzą ze złóż morskich, jedynymi wyjątkami są E. cromptonellus z przypuszczalnego złoża jeziornego oraz nowo znaleziona szczęka z lądowej Formacji Chinle.
W rzeczywistości Dalla Vecchia (2013) zauważył, że wszystkie jednoznaczne okazy noryckich pterozaurów pochodzą z tropikalnych brzegów Tetydy – prehistorycznego oceanu, który powstał jako wielka zatoka na wschodzie Pangei. Ogólny brak pterozaurów w triasowych złożach lądowych jest zagadkowy, szczególnie że istnieją miejsca triasowe ze znakomicie zachowanymi kręgowcami, takie jak Ghost Ranch, Nowy Meksyk, USA i Karoo Basin w Południowej Afryce. Jest to zagadkowe również z tego względu, że przypuszczalnie pterozaury ewoluowały od przodków lądowych.
Całkowity brak jakichkolwiek „proto-pterozaurów” w triasowych złożach lądowych stanowi problem dla naturalistycznej teorii ewolucji. Możliwym rozwiązaniem sugerowanym przez Dalla Vecchia (2013) jest to, że „proto-pterozaury” nie latały i żyły daleko od miejsc, w których miało miejsce doskonałe zachowanie kręgowców. Podobna sugestia dotyczy braku form przodków kambryjskich taksonów o twardych ciałach. Chociaż tendencyjny sposób zachowania skamielin wydaje się mieć zauważalny rezultat wśród pterozaurów (Butler i in., 2013), wydaje się, że łatwiej byłoby skamienieć nielatającym przodkom pterozaurów z ich mniej delikatnymi kośćmi niż ich latającym potomkom.
Co ciekawe, nieznane są ślady stóp pterozaurów triasowych. W rzeczywistości zapis śladu pterozaura rozpoczyna się w górnej jurze, chociaż może istnieć kilka przykładów ścieżek wydeptanych przez pterozaura z środkowej jury (Lockley i in., 2008). Nagłe pojawienie się śladów pterozaurów między górną jurą a dolną kredą zbiega się dobrze z eksplozją różnorodności gatunkowej pterozaurów oraz różnorodnością paleośrodowiskową w tym samym przedziale czasowym (McLain, 2012).
Czy potop może stanowić lepsze wyjaśnienie zapisu kopalnego pterozaura? Jest to możliwe. Ponieważ pterozaury są tak niewiarygodnie odrębne (tj. są nieciągłe w stosunku do wszystkich innych organizmów), wydaje się mało prawdopodobne, aby wyewoluowały z gadów lądowych. Zatem kreacjoniści nie spodziewaliby się, że znajdą skamieliny „proto-pterozaura”. Ponadto pterozaury mogły zostać pochowane na podstawie preferencji siedliskowych lub zdolności lotu, a nie etapu ewolucji. To by mogło wyjaśniać, dlaczego bardziej „zaawansowane rhamphorhynchoids”, takie jak Eudimorphodon, zostały pochowane wraz z bardziej „prymitywnymi rhamphorhynchoids”, takimi jak Peteinozaur i Preondaktyl. Zamiast znajdować na dole zapisu kopalnego pterozaura pień do pterozaurowego drzewa, którego naturalistyczny paleontolog ewolucjonista mógłby się spodziewać, dowody sugerują, że najniżej w zapisie kopalnym położone pterozaury są już bardzo zróżnicowane zarówno pod względem liczby gatunków, jak i liczby rodzin.
Wniosek
Nawet krótkie spojrzenie na niewielką grupę pterozaurów dostarcza fascynującego widoku na dziwny i piękny świat, który dopiero zaczynamy rozumieć. Wydaje się, że triasowe pterozaury rzucają wyzwanie oczekiwaniom wielu badaczy, podobnie jak odkrycie pierwszego okazu pterozaura w 1784 r. było wyzwaniem dla tych, którzy nigdy nie wyobrażali sobie takiego stworzenia. Pterozaury – formy triasowe, jurajskie i kredowe – stanowią dla nas doskonałą lekcję dotyczącą nauki: kiedy to, czego oczekujemy, nie pasuje do rzeczywistości, musimy cofnąć się i rzucić wyzwanie rzeczom, o których myśleliśmy, że je znamy, ponieważ jest w nich wiele założeń, a nie wszystkie założenia są właściwe.
Autor: Matthew McLain
(Aktualizacja na temat Pterozaurów triasowych znajduje się tutaj.)
Bibliografia
- Andres, B., Clark, J., and Xu, X., 2014, The earliest pterodactyloid and the origin of the group: Current Biology, doi:10.1016/j.cub.2014.03.030.
- Barrett, P.M., Butler, R.J., Edwards, N.P., and Milner, A.R., 2008, Pterosaur distribution in time and space: an atlas: Zitteliana, v. B28, p. 61–107.
- Bonaparte, J.F., Schultz, C.L., and Soares, M.B., 2010, Pterosauria from the Late Triassic of Southern Brazil, inBandyopadhyay, S. ed., New Aspects of Mesozoic Biodiversity Lecture Notes in Earth Sciences, v. 132, p. 63-71.
- Butler, R.J., Barrett, P.M., Nowbath, S., and Upchurch, P., 2009, Estimating the effects of sampling biases on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement: Paleobiology, v. 35, no. 3, p. 432–446.
- Butler, R.J., Benson, R.B.J., and Barrett, P.M., 2013, Pterosaur diversity: untangling the influence of sampling biases, Lagerstätten, and genuine biodiversity signals: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 372, p. 78–87.
- Dalla Vecchia, F.M., 2002, A caudal segment of a Late Triassic pterosaur (Diapsida, Pterosauria) from North-eastern Italy: Gortania, v. 23, p. 31-58.
- Dalla Vecchia, F.M., 2009a, Anatomy and systematics of the pterosaur Carniadactylus n. rosenfeldi(Dalla Vecchia, 1995), Rivista Italiana di Paleontologia e Stratigrafia, v. 115, no. 2, p. 159-186.
- Dalla Vecchia, F.M., 2009b, The first Italian specimen of Austriadactyluscristatus (Diapsida, Pterosauria) from the Norian (Upper Triassic) of the Carnic Prealps, Rivista Italiana di Paleontologia e Stratigrafia, v. 115, n. 3, p. 291-304.
- Dalla Vecchia, F.M., 2013., Triassic pterosaurs, inNesbitt, S.J., Desojo, J.B., and Irmis, R.B. eds., Anatomy, Phylogeny, and Palaeobiology of Early Archosaurs and their Kin, 379, Geological Society of London, Special Publications, p. 119-155.
- Dalla Vecchia, F.M., Wild, R., Hopf, H., and Reitner, J., 2002, A crested rhamphorhynchoid pterosaur from the Late Triassic of Austria: Journal of Vertebrate Paleontology, v. 22, n. 1, p. 196-199.
- Jenkins, F.A., Jr., Shubin, N.H., Gatesy, S.M., and Padian, K., 2001, A diminutive pterosaur (Pterosauria: Eudimorphodontidae) from the Greenlandic Triassic: Bulletin of the Museum of Comparative Zoology, v. 156, no. 1, p. 151–170.
- Kellner, A.W.A., 1996, Description of new material of Tapejaridae and Anhangueridae (Pterosauria, Pterodactyloidea) and discussion of pterosaur phylogeny, PhD thesis, Columbia University, [Published by University Microfilms International/ProQuest].
- Kellner, A.W.A., 2003, Pterosaur phylogeny and comments on the evolutionary history of the group, inBuffetaut, E. and Mazin, J.-M. eds., Evolution and Palaeobiology of Pterosaurs, 217, Geological Society of London, Special Publications, p. 105–137.
- Kellner, A.W.A., 2010, Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species: Anais da Academia Brasileira de Ciências, v. 82, no. 4, p. 1063–1084.
- Lockley, M., Harris, J.D., and Mitchell, L., 2008, A global overview of pterosaur ichnology: tracksite distribution in space and time: Zitteliana, v. B28, p. 185–198.
- McLain, M. A., 2012, Pterosaur diversity parallels changes in paleoenvironment, Geological Society of America Abstracts with Programs, v. 44, no. 7, p. 401.
- McLain, M.A., Chase, B., and Bryant, E., 2014, PteroTerra: a searchable pterosaur database web application that interfaces with Google Earth: Historical Biology, doi: 10.1080/08912963.2014.929671.
- Unwin, D.M., 2003, On the phylogeny and evolutionary history of pterosaurs, inBuffetaut, E. and Mazin, J.-M. eds., Evolution and Palaeobiology of Pterosaurs, 217, Geological Society of London, Special Publications, p. 139-190.
- Unwin, D.M., 2006, The Pterosaurs from Deep Time: Pi Press, New York, New York.
- Wellnhofer, P., 1991, The Illustrated Encyclopedia of Pterosaurs: Salamander Books Ltd, London, United Kingdom.
- Wellnhofer, P., 2003, A Late Triassic pterosaur from the Northern Calcareous Alps (Tyrol, Austria), inBuffetaut, E. and Mazin, J.-M. eds., Evolution and Palaeobiology of Pterosaurs, 217, Geological Society of London, Special Publications, p. 5-22.
- Whatley, R.L., Behrensmeyer, A.K., McIntire, S.B., Ramezani, J., and Parker, W.G., 2013, First pterosaur dentary and postcrania from the Upper Triassic Owl Rock Member, Chinle Formation, Petrified Forest National Park, Arizona: Journal of Vertebrate Paleontology Annual Conference Program and Abstracts,, p. 237.
- Wiffen, J., and Molnar, R.E., 1988, First pterosaur from New Zealand: Alcheringa, v. 12, p. 53–59.
